定義:上皮組織は細胞のみで構成され、これらの細胞は接合質や突起、あるいはその両方によって互いに結合している。1層または多層を成して連続し、体の内外にある自由表面を被覆している。
上皮組織は細胞のみから成るため、あらゆる組織の中で最も単純な構造を持つ。そのため、細胞学の直後に論じるのが適切である。
一般的性状:上皮細胞は原形質と核から成り、明確な境界を持つ。細胞膜は往々にして欠如し、その代わりに原形質の周縁部が固化している場合が多い。大多数の上皮細胞は柔軟で、周囲の圧力に容易に適応できる。しかし、その配列ゆえに、状況によっては互いにかなり強い圧力を及ぼし合うことができる。これは特に胎生期において、形態発生の重要な現象を引き起こす。成体においても、多くの上皮細胞群はその構成要素、すなわち個々の細胞が互いに及ぼす圧力下にある。この事実は細胞の形態から容易に推測でき、他の観点からも証明可能である。
上皮組織を構成する細胞の形態の多様性Formenreichtumは顕著であり、大きさの差異も著しい。細胞体の外面および内部には多種多様な構造が観察されることがある。
形状に基づいて4種類に分類される:扁平上皮Platten-Epithel(敷石上皮Pflaster-Epithelとも呼ばれる)、円柱上皮Zylinder-Epithel、繊毛上皮Flimmer-Epithel、移行上皮Übergangs-Epithelである。
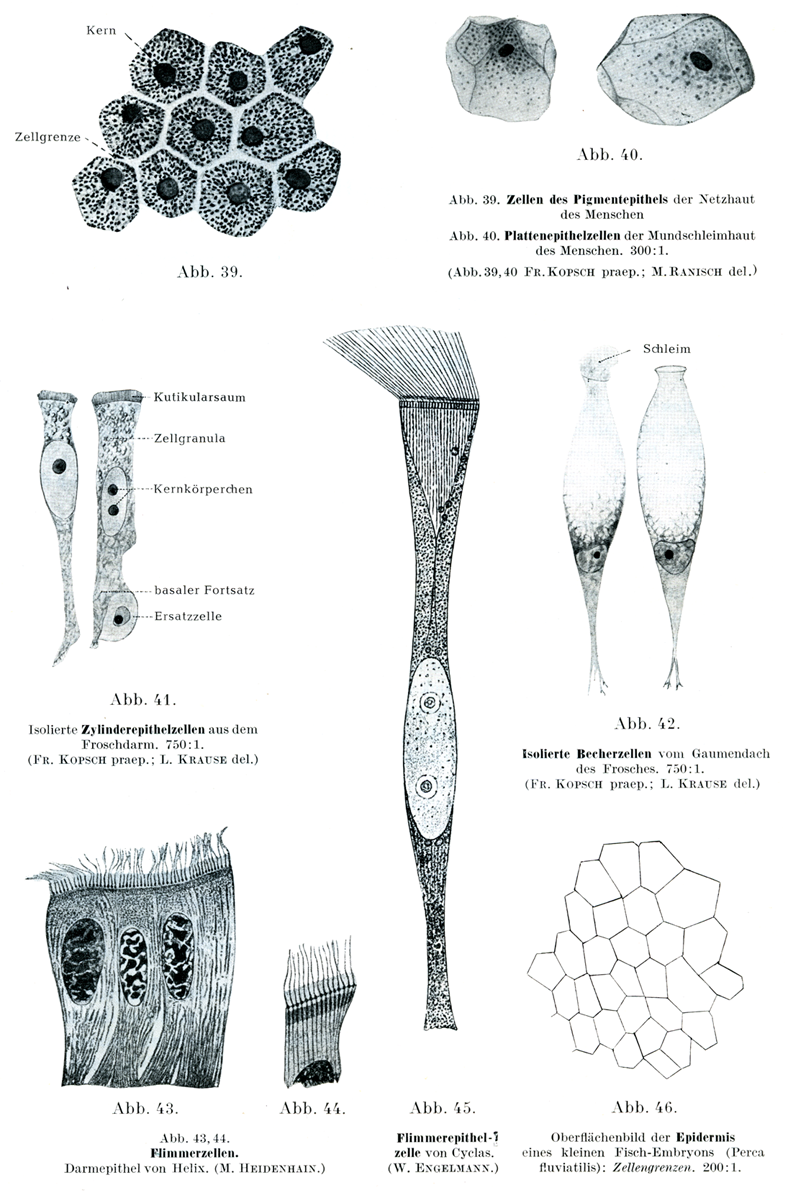
扁平上皮細胞は平坦で薄く、小鱗状の板を形成し、その境界線は多くの場合不規則である。例外として、網膜の色素上皮(色素層Stratum pigmenti)の細胞は規則正しい六角形を呈する(RK039(色素上皮細胞)、040(扁平上皮細胞)、041(円柱上皮細胞)、042(杯細胞)、043-044(絨毛細胞)、045(絨毛上皮細胞)、046(小型胚子) )。一方、口腔上皮の表面細胞は不規則な輪郭を示す(RK039(色素上皮細胞)、040(扁平上皮細胞)、041(円柱上皮細胞)、042(杯細胞)、043-044(絨毛細胞)、045(絨毛上皮細胞)、046(小型胚子) )。その他の扁平上皮細胞でも五角形あるいは六角形を呈するものがあり、その境界は直接的だが、境界線の長さは様々である(RK039(色素上皮細胞)、040(扁平上皮細胞)、041(円柱上皮細胞)、042(杯細胞)、043-044(絨毛細胞)、045(絨毛上皮細胞)、046(小型胚子) )。
円柱上皮細胞は様々な長さの角柱の形状を呈する。これには(いわゆる)立方形細胞kubische Zellen[「立方形」細胞"kubische" Zellenという名称は適切ではない。サイコロ形の細胞を誰も見たことはないだろう。いわゆる立方形細胞は、その形状から言えば五角形あるいは六角形の円柱状または角柱状の細胞で、高さの低いものである(原著註)]から、眼の水晶体を構成する長い糸状の「水晶体線維」Linsenfasernまで、あらゆる形態のものがある。楕円に近い形の核が、その長軸を細胞の縦方向に一致させて存在する。細胞の基底部は細い突起を出して、その下にある組織に鉤のように付着している。
絨毛細胞(Flimmer- oder Wimper-Zellen)は円柱状または円錐状の細胞で、その自由面に多数の細かい毛が生えている。この毛は、生細胞内で運動(bewegen od. flimmern)する。絨毛は小皮縁(Kutikularsaum)という細胞体縁の特殊な硬化層に付着し、そこで基底小体(Basalkörperchen)と結合している(RK039(色素上皮細胞)、040(扁平上皮細胞)、041(円柱上皮細胞)、042(杯細胞)、043-044(絨毛細胞)、045(絨毛上皮細胞)、046(小型胚子) )。細胞体内部には細い糸状物が伸び、これらが集まって円錐状となり、絨毛根円錐(Wimperwurzelkegel)と呼ばれる。楕円形に近い核は小皮縁からやや離れた位置にあり、その長軸は細胞の縦軸と平行である。絨毛細胞の基底部は円錐状に細まり、尖った突起を形成する。この突起は時に数本の細い糸状物に分かれ、上皮下層に鈎状に付着する。
個々の細胞を観察すると、絨毛の運動様式は多様だが、毛のなびく方向は一定している。弱拡大で小さな細胞を観察する場合、絨毛細胞の特性を識別するのは難しい。多種の動物を比較研究することで、初めて絨毛細胞の複雑な構造がより明確になる。絨毛細胞は動物界全体で重要な役割を果たし、時にはこれが唯一の運動器官となる(RK039(色素上皮細胞)、040(扁平上皮細胞)、041(円柱上皮細胞)、042(杯細胞)、043-044(絨毛細胞)、045(絨毛上皮細胞)、046(小型胚子) )。
絨毛は単層扁平上皮や様々な高さの円柱上皮にも存在し、後者は単層または重層である(RK047(ヒト気管の多列線毛上皮))。K. Peterの研究(Anat. Anz., 15. Bd., 1898, 1899)によれば、絨毛細胞の無核部分でも活発な運動が見られる。つまり、核は絨毛の動きに直接関与しない。また、原形質から分離した毛も運動することから、原形質も直接的な影響を持たないことが分かる。絨毛運動の中心は絨毛装置自体にあり、基底小体がその役割を担うと考えられる。この考えは精子(Spermien)の断片に関する所見とも一致し、中部(Mittelstück)と連続する断片のみが運動を示す。しかし、基底小体と中心小体の同一性はまだ確定されておらず、植物に関する問題も未解決である(詳細は Heidenhain, Plasma und Zelle. Bd. I., Jena, 1907, p.287 参照)。v. Renyi(Zeitschr. Anat. Entwgesch., 81. Bd., 1926)は新しい方法(生体染色、微小操作)を用いて絨毛細胞の運動中心を研究したが、「これまでの記述的・実験的形態学では、絨毛細胞の運動中心を確実に決定することはできていない」という悲観的な結論に達した。
円柱上皮細胞や絨毛上皮細胞の集団中に、機能面では粘液細胞(Schleimzellen)、形状面では杯細胞(Becherzellen)と呼ばれる特殊細胞が存在する。これらの細胞の形状は分泌物の量に応じて変化する。分泌物が少ない時は円柱細胞に似ているが、粘液が増加すると樽状になる。最初は細長い形状だが、徐々に膨らみ丸みを帯びていく。核は残存する原形質とともに細胞の基底部へ移動し、その形も変化する。細胞の自由端が開口すると、粘液が密雲や綿雪のように膨らみ、開口部から分泌される(RK039(色素上皮細胞)、040(扁平上皮細胞)、041(円柱上皮細胞)、042(杯細胞)、043-044(絨毛細胞)、045(絨毛上皮細胞)、046(小型胚子) 、RK055(角膜の上皮細胞)、056(円柱上皮)、RK057(多列繊毛上皮)、058(中心鞭毛装置) )。
閉鎖堤(Schlussleisten)は接合質の特殊な形態で、多くの上皮(特に様々な粘膜の円柱上皮や移行上皮)に見られる。これは上皮細胞の表面に向かった端を結合しており、表面から見ると閉鎖堤網(Schlussleistennetz)という網状の構造を示す。
細胞間橋については、一つの細胞から他の細胞へ続く原形質の突起、細胞膜の突起、あるいはその両方という異なる見解がある。その本質を決定するのは難しい。仮に原形質の突起だとすると(動物細胞が通常細胞膜を欠くことからこの説が有力だが)、全身の広範な細胞群の原形質が無数の突起で相互に連結しているという重要な意味を持つ。これらの突起は胎児期から存在が確認されており、細胞同士を強固に結合させる役割を早期から果たしている。
一方で、細胞間隙(Interzellularlücken)という微小な管も残存しており、そこを細胞間液(interzelluläre Flüssigkeit)、別名上皮リンパ(Epithellymphe)が流れている。この液体は上皮の栄養に極めて重要な役割を果たす。さらに、遊走細胞もこの管を通って上皮表面に到達する。
上皮層形成:すでに述べたように上皮細胞が集まって、一続きの被い、すなわち上皮を形成している。重なり合っている細胞層の数によって
a) 単層上皮 einfaches (einschichtiges) Epithel
b) 重層上皮 geschichtetes (mehrschichtiges) Epithel
が区別される。
[図39] ヒトの網膜の色素上皮細胞
[図40] ヒトの口腔粘膜の扁平上皮細胞(×300)
[図41] カエルの腸から単離した円柱上皮細胞(×750)
[図42] カエルの口蓋から単離した杯細胞(×750)
[図43・44] 絨毛細胞:ヨーロッパの食用カタツムリ(Helix)の腸上皮(M. Heidenhain)
[図45] ドブシジミ(Cyclas)の絨毛上皮細胞(W. Engelmann)
[図46] 硬骨魚スズキ属(Perca fluviatilis)の小型胚子の表皮を外側から観察:細胞境界を示す(×200)

[図47]ヒト気管の多列線毛上皮。
Kopsch-Kolatschev法により細胞内の内部骨格を可視化。1000倍

[図48]サンショウウオの腸上皮細胞における粘液形成。A:小さい粘液顆粒をもつ細胞。B:大きい粘液顆粒をもつ細胞。(Heidenhain, Plasma und Zelle. Bd. Iより)
[図49]家兎の臍胱から得た移行上皮細胞。×300倍。左上:表層の幅広い扁平細胞。2つの核を有し、下面に鋭く突出した明瞭な縁とくぼみがある。下:より深層に属する2つの西洋梨型細胞。右上:1個の西洋梨型細胞が、表層細胞のくぼみにはまり込んでいる。(KleinのデータをもとにSchäferが描画)

[図50]尿管の移行上皮 ヒト尿管粘膜の横断図。500倍拡大。

[図51]閉鎖堤網の模式図(Stöhr sen. による)
[図52]細胞間橋 ヒト表皮の切片(×1000倍)。細胞間隙が拡大しており、そのため細胞間橋にランヴィエ小節(Ranviersche Knötchen)が見られない。*は核小体を示す。
理論的には、この二つの型は上述の4種の上皮細胞のいずれにも存在しうるはずであるが、単層の移行上皮は存在しない。したがって、以下の7種の上皮を観察することができる。
特筆すべきは、最表層のみが扁平な細胞で構成される点である。最下層の細胞は円柱状で、その上に多角形の細胞が積み重なる。これらの細胞は隣接する細胞からの圧力により多角形となる。表面に近づくにつれ細胞は扁平化し、最表層では薄い小鱗状を呈する。細胞の形状変化に伴い、核の形態と位置も変化する。下層の円柱細胞では核はほぼ楕円形、中間層では球形に近く、上層では扁平となる。
上皮細胞の表面および内部に見られる分化(äußere und innere Differenzierungen)は極めて多様である。その全容を詳述することは冗長になるため、ここではその豊かな多様性の概要を簡潔に述べるにとどめる。
上皮細胞の表面の分化として、既に細胞間橋と繊毛について言及した。運動性の付属物である運動毛(Kinocilien)に加え、細胞の自由表面に非運動性の不動毛(Stereocilien)が存在することがある。また、精巣上体管などの細胞表面には細い毛の束が見られ、これは分泌物の輸送に関与している(第II巻参照)。
中心鞭毛装置(Zentralgeißelapparat)は、細胞の自由表面から突出する短い1本の精細な毛(鞭毛Geißelまたは外糸Außenfaden)、その直下の双心子(Diplosoma)形態の中心小体、そして中心小体から細胞内部へ束状に伸びる細い糸から構成される。この鞭毛の運動性は未だ明確ではないが、形態学的には精子細胞の軸糸形成初期段階に類似している。しかし、感覚器である可能性が高く、あるいは退化した構造物かもしれない。ヤツメウナギの原腎細胞に比較的強靭な鞭毛が1本ずつ観察されることがその根拠となる。